
Dans un article du Bulletin des armées M. Edmond Perrier a fait très justement remarquer combien les périodes d’immobilité sur front se prêtent à l’étude de la nature. L’observateur attentif y trouve, en effet, mille occasions de voir et d’expérimenter. Dans le domaine végétal, laboratoire toujours à portée, il est particulièrement facile et intéressant d’étudier les effets de la lumière sur les plantes, et les moyens si ingénieux qu’elles ont de recevoir et d’absorber les rayons solaires.

Des observations journalières et des expériences très simples (celles qui sont ici rapportées ont été faites pour la plupart en campagne 1915-1916) peuvent conduire à des données nouvelles et utiles, applicables notamment à la cure solaire qui est un moyen très efficace de guérison des malades et des blessés.
La plante puise dans le rayonnement solaire une grande part de son énergie et tend vraisemblablement pour cette raison à se faire au soleil la plus large place possible. Elle s’étale en feuilles minces et vertes (le vert presque autant que le noir absorbe la lumière) ( [1]) sur une surface extraordinairement grande relativement à sa masse totale.
Voici, par exemple, un figuier de 5 m. 50 de hauteur ; il porte environ 2150 feuilles, faisant une surface totale d’environ 49 mètres carrés pour un poids de feuilles de 17 kg. Un tiers environ de cette surface baigne clans la lumière solaire directe et le reste dans la lumière diffuse. Les feuilles s’étagent et s’étalent, leur face supérieure vernissée, la plus verte, regardant la lumière. D’après des expériences antérieures ( [2]) nous avons pu calculer que ce figuier absorbe en une journée d’été plus de 40.000 grandes calories de rayonnement solaire.
La plus grande partie de cette énergie (environ 32 000 calories) est dépensée par l’évaporation d’environ 45 ( [3]) litres d’eau qui provoque dans les racines un appel proportionnel d’eau chargée de sels. Le reste sert vraisemblablement aux fonctions vitales, à la statique de la plante et à l’élaboration de la matière vivante et de ses réserves.
Toutes sortes de procédés sont bons à la plante pour se mettre au soleil : courbures, allongement et croissance régulière ou inégale, torsion ou rotation des tiges et des pétioles, flexion, redressement, retournement des limbes , mouvements variables suivant les circonstances mais toujours adaptés au but qui est d’absorber par la face supérieure des feuilles le maximum possible de radiations.
A l’air libre, s’il n’y a nul obstacle, la plante s’oriente vers le ciel et au midi ; certaines (tournesol) suivent même le soleil dans son déplacement journalier. Si l’éclairage est unilatéral, la plante se tourne toujours du côté d’où vient le plus de lumière, quelle que soit d’ailleurs l’orientation de ce côté ; elle sait pour cela et juste dans la mesure utile se courber, se dresser, se retourner, s’allonger.

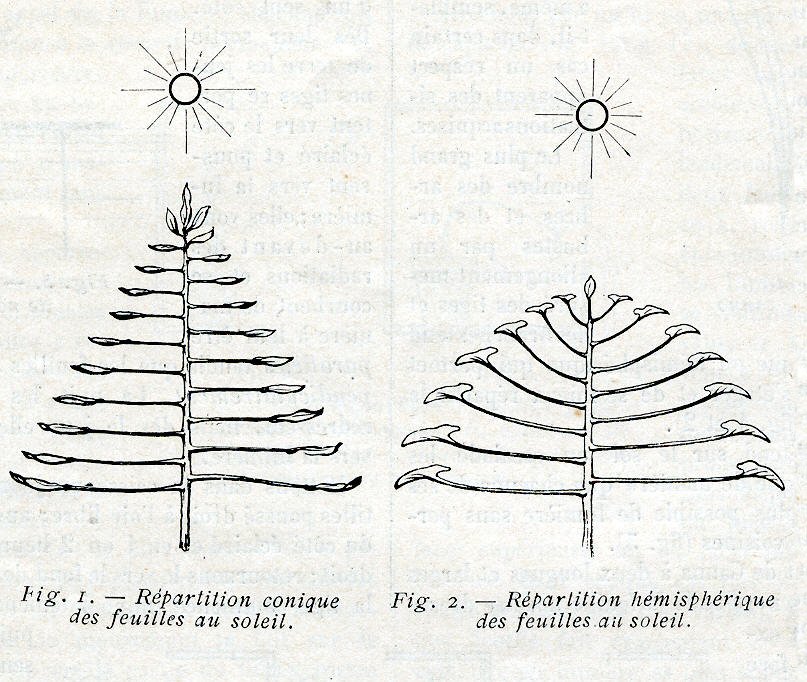
Il y a dans l’ensemble des feuilles d’une même plante, et aussi, semble- t-il, des plantes de même espèce vivant sur un même terrain, une coordination manifeste qui paraît correspondre à une volonté commune, à une entente pour le meilleur rendement total. Il y a même, semble-t-il, dans certain cas un respect apparent des situations acquises. Le plus grand nombre des arbres et des arbustes par un allongement mesuré des tiges et des branches tend à la forme conique ou hémisphérique qui permet aux feuilles de s’étager et de se mieux répartir la place au soleil (fig. 1 et 2).
Le lierre s’étend sur le sol ou escalade les murs et s’imbrique de manière que chacune de ses feuilles ait le plus possible de lumière sans porter ombre à ses voisines (fig. 3).
Voici un plant de Canna à deux longues et larges feuilles orientées nord-sud ; la feuille nord se dresse verticalement et expose au midi sa face supérieure vernissée ; la feuille sud pareillement dressée, regarderait le nord et gênerait sa voisine ; elle évite ce double écueil en se retournant et s’inclinant vers le sol (fig. 4).
Sur l’étang les feuilles de nénuphar se répartissent l’espace disponible : les plus anciennes reposent à la surface de l’eau, absorbant à la fois air, eau et soleil ; les feuilles plus jeunes sont encore immergées, ou bien se haussent quand la place est prise, entre deux feuilles anciennes et se maintiennent à demi fermées au-dessus de l’eau dans une attitude qui réduit l’ombre portée (fig. 5).

Le sens ou si l’on préfère l’instinct phototropique des plantes, visible dans leurs attitudes naturelles, apparaît plus encore dans des dispositifs d’expériences qui les obligent à se déplacer, à faire sous nos yeux pour atteindre leur but des mouvements relativement étendus et rapides.
Plaçons un semis de lentilles dans une caisse fermée par le haut et ouverte d’un seul côté. Dès leur sortie de terre les jeunes tiges se portent vers le côté éclairé et poussent vers la lumière ; elles vont au-devant des radiations et se courbent de manière à leur être parallèles tandis que les feuilles se disposent perpendiculairement. La nuit les jeunes tiges se redressent, mais dès le jour elles se recourbent vers la lumière.
Mettons dans la caisse un jeune plant de lentilles poussé droit à l’air libre ; aussitôt il se courbe du côté éclairé et en 1 ou 2 heures atteint l’angle droit ; retournons-le vers le fond de la caisse ( fig. 6) ; la tige commence immédiatement à se redresser, puis se courbe en sens inverse ; en une heure elle est droite ; en 2 ou 3 elle est de nouveau courbée vers la lumière. En 120 ou 150 minutes le sommet de la tige a fait dans un plan vertical un mouvement de 150 à 180°.
Tournons maintenant le pot de 90° seulement (fig. 7), le sommet de la tige vers un des côtés de la caisse : la plante cette fois ne se redresse pas, son extrémité supérieure reste horizontale et, sans s’incurver en avant, se tourne vers la lumière par un mouvement de rotation dans un plan horizontal sur la partie inférieure de la tige qui sert de pivot ; elle prend ainsi le plus court chemin pour revenir à la lumière.
Les tiges font leurs mouvements et leurs courbures phototropiques le plus souvent sur leur partie supérieure, la partie inférieure restant verticale quand la tige est courte et résistante ; mais quand elle est longue et grêle, il se produit souvent sur cette partie inférieure une courbure inverse, une courbure de compensation qui permet à la plante de résister à la pesanteur (fig. 8).
Les feuilles par le dispositif des tiges et par leur répartition phyllotaxique se trouvent orientées vers la lumière ; elles répondent aussi directement individuellement à l’appel de la lumière ; chacune sait se placer de manière à la recevoir normalement et par sa face supérieure, chacune s’arrange au besoin et fait son mouvement pour son compte, à sa manière et généralement sans gêner ses voisines.

Voici dans un appartement, un pot de géranium ; toutes les tiges terminales et toutes les feuilles « regardent » la fenêtre ; retournons-le face au mur ; en deux ou trois jours toutes les tiges et toutes les feuilles font demi-tour et face à la lumière ( [4]).
Le mouvement se fait le plus souvent sur le pé- tiole qui se courbe et se déplace exactement comme les tiges ; mais si un obstacle s’oppose à la courbure des pétioles, le mouvement se fait sur le limbe lui-même ou sur la partie du limbe restée libre Retournons et attachons contre un mur une feuille de géranium grimpant, le limbe vertical et dos à la lumière ; en 2 ou 3 jours, la feuille se retourne en pivotant sur le pétiole.

Des feuilles de vigne (fig. 10) étroitement attachées dans ces conditions contre la treille ont mis 15 à 20 jours pour se retourner complètement ; mais leur mouvement a été plus complexe ; les 3 premiers jours, la feuille collée au mur a relevé ses bords, puis elle a commencé sur le pétiole un double mouvement de rotation d’arrière en avant et de torsion latérale. Avec du liège et deux épingles, fixons par sa base sur le bord d’une planche horizontalement et la face supérieure vers le sol une feuille de fraisier (fig. 11). En 3 ou 4 jours, souvent moins, les 3 lobes de la feuille, chacun séparément et de manière différente, se redressent ou se retournent vers la lumière, soit par une courbure du limbe, soit par une torsion du pétiole dont les fibres superficielles prennent un aspect spiralé.

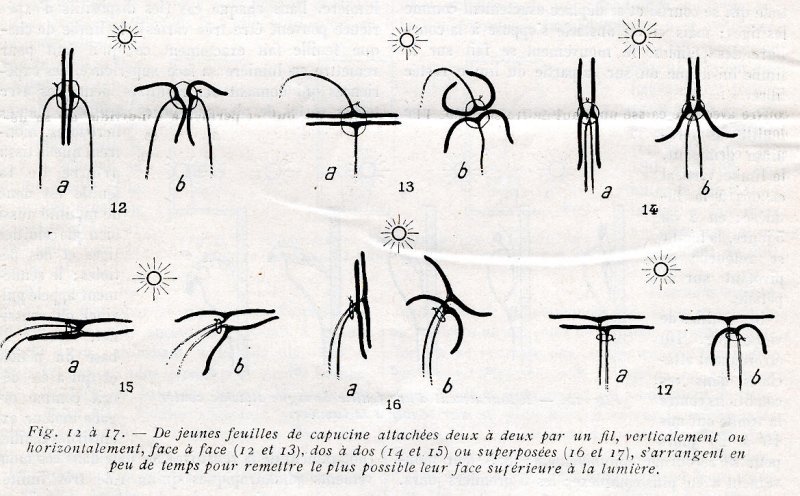
Attachons par un fil ou une épingle deux feuilles de capucine (fig. 12 à 17), de manière que leurs faces supérieures soient accolées verticalement ou horizontalement ; dès le lendemain, les bords des deux feuilles sont écartés en X, relevés ou abaissés, et la lumière pénètre entre les limbes. L’écartement se continue les jours suivants et les limbes arrivent à se retourner, à se replier complètement. Des mouvements analogues se produisent si les feuilles sont disposées dos à dos, ou bien superposées les faces supérieures ou les faces inférieures vers la lumière. Dans chaque cas (les dispositifs d’expérience peuvent être très variés), le limbe de chaque feuille fait exactement ce qu’il faut pour remettre en lumière sa face supérieure. Ces expériences qui donnent des résultats identiques avec toutes .sortes de plantes, arbres, arbustes, plantes herbacées, montrent que le tissu propre de la feuille est doué de motilité aussi bien que celui des tiges et des pétioles ; le renflement appelé pulvinel ou coussinet, situé à la base du pétiole et qui a été décrit comme organe moteur exclusif des feuilles (notamment de la sensitive) ne joue dans ces mouvements phototropiques qu’un rôle très limité ; toutes les parties souples des plantes paraissent d’ailleurs capables de mouvements.

Les fleurs subissent, elles aussi, la loi héliotropique et veulent recevoir la lumière normalement et pour la plupart sur leur face supérieure. Elles aussi s’étagent, s’étalent, se haussent sur leur tige, s’inclinent ou se redressent vers la lumière. Comme les feuilles, elles font des mouvements étendus et relativement rapides pour retrouver la lumière si on les en détourne.
Attachons tête en bas par un fil, une fleur de zinia, de capucine, de reine-marguerite (fig. 18) ; en quelques heures la fleur se redresse ( [5]) par une courbure de la tige. Le mouvement est aisé et rapide si l’attache est à une certaine distance de la fleur, plus lent et parfois incomplet si l’attache est juste à sa base.
Voici un cas la fleur et qui montre son habileté et la complexité de son effort vers la lumière : couvrons d’un capuchon de papier noir convenablement mais pas trop étroitement attaché, la moitié ou deux tiers d’une fleur de zinia (fig. 19) ; en quelques jours la fleur parvient à se désencapuchonner.

A quelques variantes près, les résultats de ces expériences sur les tiges, les feuilles et les fleurs sont constants et peuvent être obtenus avec des espèces et des plants différents et plusieurs fois de suite sur un même sujet. La plante obéit chaque fois à l’incitation de la lumière. Cependant après plusieurs épreuves rapprochées et de sens différents, les mouvements sont plus lents, la plante paraît fatiguée.
Si on expose sous une cloche de verre à des vapeurs d’éther de jeunes plants de lentilles, ceux-ci restent ensuite plusieurs heures (12 à 20 heures) sans obéir au phototropisme ; mais il n’est pas certain qu’il s’agisse là d’une véritable anesthésie ; le phototropisme exige, en effet, pour se manifester normalement une vitalité normale. Les sujets malingres, malades ou meurtris, obéissent mal. Les jeunes plants très souples sont particulièrement sensibles et mobiles. A un certain âge, les tiges trop rigides ne s’incurvent plus, mais les feuilles et les fleurs quel que soit l’âge de la plante se tournent presque toujours vers la lumière en un temps plus ou moins long.

Parfois, mais très rarement, la plante fatiguée ou maladroite ne réussit pas son mouvement. Nous avons vu une feuille de poirier, mourir en se retournant au soleil par torsion sur le pétiole. La torsion a été tellement accentuée que la circulation s’est arrêtée brusquement le 3ejour, la feuille s’était étranglée de son propre mouvement.
La rapidité du phototropisme ne dépend pas de l’intensité de la lumière. Au contraire, pour les jeunes tiges, le rayonnement diffus résultant d’un éclairage intense paraît retarder les mouvements ; ceux-ci sont d’autant plus rapides et accusés que l’éclairage est plus unilatéral. En chambre noire, la lumière d’une lampe ou d’une bougie le provoque aussi vite que la lumière du jour. Mettons un jeune plant de lentille dans une caisse complètement fermée, mais ayant sur un de ses côtés une petite fenêtre, et près de celle-ci, au dehors, allumons une bougie ; en deux ou trois heures, la tige se courbe complètement et les feuilles terminales viennent se placer au centre de la fenêtre (fig. 9). On sait d’ailleurs quelles longueurs de tige et quelles courbures extraordinaires présentent les plantes en cave pour arriver à la lumière. « Lorsqu’un végétal est privé de lumière, il allonge « ses tiges comme s’il cherchait quelque issue pour « retrouver la lumière qui lui manque »( [6]) Il semble que l’effort de la plante est d’autant plus accusé que son appétit de lumière est moins satisfait.
Un autre fait très remarquable est que les mouvements phototropiques commencés le jour se continuent la nuit et que le retournement des feuilles et des fleurs peut même se faire entièrement dans l’obscurité. En effet si l’on couvre avec une caisse un plant de fraisier (fig. 11) dont une feuille a été retournée vers le sol, celle-ci se redresse quand même en deux ou trois jours dans la direction d’où lui venait la lumière.
Comment expliquer ces diverses manifestations du phototropisme des plantes ? Les anciens naturalistes tendaient à voir dans les attitudes héliotropiques des feuilles et des fleurs une orientation instinctive de la plante. Dans la suite on a cherché à les expliquer par des causes purement physiques.
Darwin, qui a particulièrement étudié les mouvements dits de circumnutation des tiges et des racines en voie de développement, les a comparés aux mouvements inconscients des animaux inférieurs ; les mouvements héliotropiques des tiges et des feuilles dériveraient, par adaptation à un but utile, de la circumnutation et seraient « régularisés mais non directement déterminés par la direction des rayons lumineux&nbp; ».
Pour d’autres auteurs les courbures phototropiques résulteraient d’une action mécanique directe, d’une sorte de pression des rayons lumineux sur les tiges et les pétioles ou d’une modification de la turgescence et de la croissance du côté éclairé. Cette théorie mécanique du phototropisme particulièrement soutenue par les Allemands était en dernier lieu la plus généralement acceptée.
Les observations et expériences précédentes montrent, croyons-nous, qu’il y a dans le phototropisme des plantes, comme dans celui des animaux, non un phénomène mécanique passif, ni simplement une modification de la croissance due à la lumière, mais la manifestation d’une sensibilité et d’une motilité ( [7]), qui par ailleurs se révèlent de bien des manières.
Il n’est pas exagéré, croyons-nous, de voir aus dans le phototropisme végétal la manifestation d’une fonction directrice, intelligente ( [8]). ) en ce sens qu’elle préside dans les différents cas à l’adaptation des moyens au but et permet, à l’individu de se défendre et d’atteindre, malgré les obstacles les plus divers, ce dont il a besoin. Comme les animaux, les plantes cherchent leurs aliments l’air, l’eau, le soleil, savent aller au-devant d’eux et les trouver, faisant preuve de sensibilité spéciale et d’un admirable sens de direction. A les regarder de près, elles apparaissent comme des êtres très perfectionnés, assez proches de nous, supérieurs même à certains points de vue biologiques. Le médecin et l’hygiéniste trouvent en elles plus d’un exemple à suivre dans l’art de vivre et de guérir et doivent particulièrement étudier et chercher à imiter leur merveilleuse manière d’utiliser l’énergie solaire.
