Atropine et Pilocarpine
Les deux poisons que je prends ici comme types de substances antagonistes ont été déjà très étudiés, soit au point de vue des effets isolés propres à chacun d’eux, soit au point de vue de leur action tant antagoniste qu’antidotique [1]. Aux faits déjà connus, j’ajouterai quelques faits nouveaux qui me paraissent de nature à éclairer leur mode d’action physiologique ; d’autre part, j’essayerai d’établir, entre ces données éparses, un lien théorique qui (autant que cela est possible en physiologie) permette de les comprendre dans quelques formules générales et simples.
I.
Je suppose connus les effets communément indiqués de l’une et de l’autre substance. On injecte, par exemple, dans le tissu cellulaire d’un chien quelques milligrammes d’un sel neutre d’atropine en solution dans l’eau. Dès que la substance est absorbée et qu’elle circule dans le sang, on voit la pupille se dilater, le cœur accélérer ses battements, les sécrétions se tarir. Ces trois effets parallèles, simultanés, paraissent au premier abord de nature assez dissemblable ; mais ce n’est là qu’une apparence. Ces trois organes, l’iris, le cœur, les glandes, sont, au contraire, pour le physiologiste, très semblables entre eux : ils appartiennent à une catégorie commune d’appareils dont le groupement constitue, par opposition avec un autre ensemble d’organes, l’une des divisions fondamentales de la physiologie. Ajoutons tout de suite que l’atropine n’agit pas sur ces appareils eux-mêmes, mais sur les nerfs qui les commandent, et le lien qui existe entre ces trois effets disparates apparaît déjà plus clairement ; pour être plus précis, ne disons pas seulement qu’elle agit d’une façon quelconque, mais qu’elle paralyse ces nerfs de la même manière que le curare paralyse certains autres nerfs moteurs (les nerfs moteurs volontaires) : l’atropine est une sorte de curare des nerfs de la vie involontaire ou inorganique.
La preuve expérimentale de celte action curarisante est facile à donner, au moins pour certains nerfs. Sur l’animal atropinisé, les nerfs sécréteurs sont inexcitables, et c’est même là une donnée banale en physiologie. De même pour la pupille : sa dilatation provient du défaut d’activité du sphincter irien ; mais là encore, c’est le nerf seul qui est inexcitable, le muscle est simplement inexcité, et il se relâche. Cette inexcitabilité des nerfs moteurs de l’iris était jusqu’ici un fait plutôt supposé que démontré que d’hypothèses n’a-t-on pas faites pour expliquer l’action de la belladone sur la pupille ! Il importait d’être bien fixé sur ce point. Avec M. Doyon, nous avons fait plusieurs fois l’expérience suivante : sur un chien on met à découvert le nerf oculo-moteur commun et, si l’on veut, en même temps ceux des nerfs ciliaires qui lui font suite. Par une excitation préalable, on vérifie d’abord que ce nerf a bien son excitabilité en tant que nerf moteur de l’iris. On injecte alors dans le sang deux milligrammes du sel d’atropine, ou plus simplement on verse dans l’œil quelques gouttes de sa solution. Après quelque temps, excite-t-on de nouveau l’oculo-moteur ou les nerfs ciliaires, la pupille reste largement dilatée, et quelle que soit l’intensité du courant excitateur, elle ne se resserre plus. Le nerf est évidemment paralysé, mais point le muscle ; car si on porte l’excitation directement sur lui, on voit alors la pupille se resserrer. Ainsi la dilatation pupillaire qui suit l’instillation de la belladone ou de son alcaloïde dans les yeux est due à l’action curarisante de ce poison sur les fibres de I’oculo-moteur commun qui commandent la contraction du sphincter de la pupille. L’atropine a de plus un effet parallèle sur l’accommodation qui s’explique de même. Comme le sphincter irien, le muscle ciliaire (muscle de l’accommodation) est relâché pendant l’empoisonnement atropique, ce relâchement est également dû à la paralysie de l’oculo-moteur commun. C’est ce dont nous nous sommes assurés également avec M. Doyen en fixant la deuxième image de Purkinje pendant qu’on excitait le nerf comme plus haut, soit avant, soit après l’instillation d’atropine dans l’œil ; avant l’action du poison, cette image diminue d’étendue par suite du bombement du cristallin résultant de la contraction du muscle ciliaire ; après, elle reste invariable quelle que soit la force de l’excitant. Ces effets paralysants disparaissent peu à peu, et le nerf reprend son excitabilité normale à mesure que le poison s’élimine.
Voyons maintenant l’effet sur le cœur ; c’est encore un effet de paralysie : à vrai dire, cette paralysie se traduit d’une façon singulière par l’accélération des battements du cœur. Mais le nerf, qui est ici paralysé, n’est pas un nerf construit sur le type ordinaire ; c’est un frein, un nerf d’arrêt, un nerf inhibiteur, suivant l’expression proposée par M. Brown-Séquard, et qui décidément doit prévaloir. Le fait de la paralysie du pneumogastrique, nerf inhibiteur du cœur, est, lui aussi, une donnée banale acceptée sans conteste par tous les physiologistes. Toutefois, le parallélisme si étroit que nous avions jusqu’ici constaté entre les effets de la belladone sur les divers appareils nerveux considérés (glande, pupille, accommodation) s’en trouve quand même altéré. L’action est bien encore paralysante, mais pourquoi s’adresse-t-elle ici aux nerfs moteurs (œil, glandes), et là aux nerfs inhibiteurs (cœur) ? Certains faits que je ferai connaître plus bas aideront, je crois, à le comprendre, et ramèneront à l’unité ces effets en apparence disparates.
II.
Nous devons maintenant porter notre attention sur les effets du poison antagoniste. À un chien nous faisons absorber une certaine quantité de pilocarpine (alcaloïde du Jaborandi), quelques centigrammes, plus ou moins, suivant la taille de l’animal. Les effets visibles, au bout d’un moment, sont bien exactement antagonistes des précédents. Les glandes sécrètent abondamment. La pupille se rétrécit, mais à la condition d’avoir instillé la substance dans l’œil ; son injection dans le sang n’a pas le même effet : l’accommodation est modifiée, elle aussi, dans un sens exactement opposé aux effets de la belladone. Le cœur se ralentit et peut même s’arrêter quand la dose est trop forte ou trop rapidement absorbée. La pilocarpine paraît donc faire exactement l’inverse de l’atropine ; celle-ci, disions-nous, est un curare, j’entends un curare d’ordre spécial s’adressant à des nerfs d’une classe particulière. La pilocarpine serait donc inversement un excitant, une sorte de strychnine de ces mêmes appareils nerveux. On l’a dit ; on paraît tout au moins l’admettre implicitement. Il me semble, pour ma part, impossible d’adopter une pareille explication : elle est contraire aux faits. La pilocarpine, elle aussi, est un curare ; c’est ce que je pense démontrer. Il nous faut pour cela analyser avec soin son action sur le cœur.
D’où vient le ralentissement du cœur ? Pour pouvoir répondre à cette question, il importe de bien se rappeler que le cœur est commandé par deux ordres de nerfs, eux-mêmes réciproquement antagonistes : les uns modérateurs ou inhibiteurs, comme il vient d’être dit, ils sont contenus dans le pneumogastrique, les autres moteurs ou excitateurs de ses mouvements, ils proviennent, en grande partie, de la chaîne du sympathique. Toute modification en plus ou en moins du nombre de ses battements pourra reconnaître pour cause ou la diminution d’action de l’un des deux ordres de nerfs, ou l’exagération de l’action des nerfs antagonistes, ou plus vraisemblablement, dans la réalité, un effet moyen résultant de l’action inégale bien qu’opposée des deux puissances contraires. Ce sera à l’expérimentation à nous dire ce qu’il en est exactement ; mais nous ne pouvons nous dispenser d’examiner ces différentes hypothèses.
Dans le cas donné, pour juger la question, j’ai fait l’expérience suivante publiée, il y a déjà plusieurs années, dans les comptes rendus de la Société de biologie.
Sur un chien, on découvre la portion de la chaîne du sympathique qui contient les nerfs accélérateurs du cœur, on excite ces nerfs et on constate, comme c’est connu, une augmentation du nombre des battements cardiaques. On soumet l’animal à l’action de la pilocarpine, et on excite de nouveau ces mêmes nerfs ; on ne constate plus d’accélération. Il est clair par là que la pilocarpine a diminué, supprimé même l’excitabilité des nerfs en question. Ce résultat nous explique déjà suffisamment le mode d’action du poison employé, et il le range évidemment dans la classe des poisons paralysant les nerfs moteurs : c’est un curare des nerfs moteurs cœur. Mais est-ce là toute son action sur le système nerveux cardiaque ? Nullement. L’effet produit par cet agent est moins localisé, plus général que ne semblerait l’indiquer notre expérience encore incomplète ; en un mot, ce n’est pas un effet spécifique, c’est un effet moyen résultant de deux effets inégaux produits par le poison sur les deux systèmes antagonistes. En effet, sur le même animal ainsi intoxiqué par la pilocarpine, découvrons le nerf pneumogastrique qui représente l’autre système, le système frénateur ou inhibiteur, et excitons-le avec nos courants. Son excitabilité devrait avoir augmenté, au moins en apparence, puisque le système antagoniste a perdu de son activité ; or on trouve, au contraire, que cette excitabilité a diminué, parfois disparu si la dose du poison a été un peu massive et, pendant quelques instants, l’excitation du pneumogastrique chez l’animal pilocarpiné n’arrête plus le cœur.
Il faut en conclure que la pilocarpine n’est pas un poison indifférent pour les nerfs inhibiteurs du cœur ; elle est pour eux aussi un curare ; mais pour une raison qui reste, celle-là, inexpliquée, tout en les atteignant, elle limite davantage son action sur eux, et, en fait, les choses se passent comme si les accélérateurs seuls étaient paralysés. En d’autres termes : l’antagonisme n’est pas entre les substances elles-mêmes, mais entre les deux systèmes de nerfs qui commandent au cœur ; et il y a tout lieu de croire que cette formule s’applique à tous les systèmes du même genre, à tous les faits d’antagonisme dit bilatéral ou réversible, comme celui qui existe entre l’atropine et la pilocarpine.
L’expérience ci-dessus me paraît, à ce sujet, fondamentale. J’ai essayé de la développer, de la compléter. Revenons à l’atropine : elle est, disons-nous, un curare des nerfs inhibiteurs du cœur ; elle paralyse le pneumogastrique en tant que modérateur cardiaque. La constatation cent et mille fois répétée de cette action a convaincu les physiologistes que c’était là toute l’explication de l’accélération des battements cardiaques qui suit son administration. La vérité est là encore, qu’à cet effet prédominant il s’en ajoute un autre nécessairement masqué par lui, puisqu’il est d’ordre inverse, et qu’on ne peut mettre en évidence que par un artifice comme plus haut. Cet artifice sera l’excitation par les courants électriques des nerfs accélérateurs chez l’animal atropinisé. Je me suis convaincu que l’atropine diminue l’excitabilité des accélérateurs ; son effet habituel d’accélération est encore un effet moyen résultant d’une différence entre deux effets opposés. Rapppelons encore un fait que les physiologistes perdent trop de vue, s’ils ne l’ignorent, c’est l’inversion d’effets de ces substances dans certains cas, chez certains animaux, avec certaines doses. L’atropine ralentit le cœur de la grenouille ; à très haute dose, elle ferait de même chez les mammifères (Meuriot).
III.
Nous devons chercher à poursuivre cette analyse, et l’appliquer aux autres organes qui subissent l’action antagoniste des deux substances : tous ne s’y prêtent pas aussi commodément que le cœur, mais, en tout cas, dans aucun je n’ai observé quoi que ce soit pouvant laisser soupçonner que les deux substances aient une action réellement inverse l’une de l’autre, et qu’elles agissent autrement qu’en paralysant (je répète à dessein en curarisant) l’une ou l’autre espèce de nerfs antagonistes dont ils sont pourvus.
Ceci implique, par conséquent, que tous les organes en question sont pourvus des deux espèces de nerfs, ou pour mieux dire, comme on ne doute pas qu’ils soient pourvus de nerfs moteurs, ceci implique qu’en plus de ces derniers, ils possèdent tous aussi des nerfs inhibiteurs.
Il y a seulement quelques années, une telle proposition eût été acceptée avec défiance, et c’est ce qui m’a fait différer celte publication dont les faits essentiels remontent à près de dix ans. Les nerfs d’arrêt des glandes, notamment, n’étaient admis que sous les plus expresses réserves. Aujourd’hui, on trouverait peu de physiologistes disposés à nier catégoriquement leur existence. Cela tient à deux choses : d’abord la question de l’inhibition gagne du terrain tous les jours ; en second lieu, beaucoup de faits nouveaux particuliers aux glandes elles-mêmes nous montrent qu’ils doivent être pourvus de nerfs de celte sorte. (Voy. Isaac Ott, Journal of Physiology ; Gley et Arloing, Arch. de physiologie. )
J’admets, pour ma part, non seulement que les glandes possèdent des nerfs inhibiteurs, mais j’admets, en outre, que la pilocarpine et les poisons du même ordre (ésérine, muscarine) paralysent ces nerfs, et que c’est là le mécanisme intime de l’effet en apparence excitant de ces substances. Elles sont les curares des nerfs d’arrêt des glandes, comme l’atropine est le curare des nerfs d’arrêt du cœur. Une fois de plus, l’antagonisme n’est pas entre les substances elles-mêmes, mais entre les deux portions du double système qui régit les glandes comme il régit l’activité de tous les organes du même ordre.
L’assimilation que je fais de ces substances à des substances de l’ordre du curare qui, entre les deux grandes divisions du système nerveux moteur (volontaire et involontaire), sauraient distinguer l’une des doux, comme les curares eux-mêmes dans leur ensemble savent distinguer les nerfs moteurs des sensitifs, cette assimilation, dis-je, je désire la justifier à divers points de vue. Après avoir cherché à analyser leur mode d’action, étudions le mode de pénétration de ces poisons dans les nerfs. Nos expériences se sont bornées, à cet égard, à calquer celles de Cl. Bernard sur le curare lui-même.
IV.
Sur un chat ou un jeune chien (au lieu de la grenouille, il faut employer ici les mammifères), on isole le nerf tibial postérieur près du jarret, on le soulève sans le couper, et après avoir lié les vaisseaux on désarticule au-dessous de lui l’articulation tibio-tarsienne, en ne laissant à la patte postérieure d’autre communication avec le tronc que par ce nerf. On injecte alors dans une veine du tronc deux centigrammes de pilocarpine en solution dans l’eau. On voit bientôt les trois pattes autres que la patte ainsi séparée se couvrir de gouttes de sueur qui perlent sur les pulpes digitales aux orifices des glandes sudoripares. Pourquoi la pilocarpine, qui baigne pourtant les origines des nerfs sécréteurs de ce membre comme celles de tous les autres nerfs, n’agit-elle pas sur eux ? C’est sans aucun doute qu’elle ne leur a pas été présentée par la voie qui convient, c’est-à-dire par la périphérie ; le curare, on le sait bien, exige, lui aussi, cette condition : il doit pénétrer par la terminaison des nerfs moteurs, jamais par leurs origines ou leur trajet. On objectera peut-être que le défaut de sécrétion est dû à l’absence de sang et de circulation dans le membre séparé ou ligaturé. Cette objection n’est pas valable ; la contre-épreuve suivante le prouve. Excitons le nerf de la patte exsangue et la sudation y apparaîtra malgré l’absence de sang et de circulation dans le membre ; l’inexcitabilité de l’appareil sudoral ne surviendra qu’après un certain délai.
La même démonstration est applicable à l’atropine : injectée dans une veine du tronc, elle paralyse les nerfs sécréteurs partout où elle peut atteindre leurs terminaisons ; elle respecte au contraire ceux que le courant sanguin n’atteint plus ; et malgré la condition en réalité défavorable créée à la fonction sécrétoire par l’anémie du membre, si on excite comparativement ses nerfs et ceux des membres non anémiés, c’est lui qui sécrète : partout ailleurs, les nerfs sont inexcitables [2].
Veut-on un autre exemple du même genre ? On s’adressera à la pupille. Une goutte de la solution de sulfate d’atropine au centième instillée dans l’œil amène, comme chacun sait, au bout d’un moment la dilatation pupillaire, et nous avons vu plus haut que le relâchement du sphincter irien provient de la paralysie du nerf qui l’anime (l’oculo-moteur commun). Dans ce cas, l’action de la belladone est purement locale, c’est évident, il lui a suffi d’atteindre les terminaisons de ce nerf pour anéantir passagèrement ses propriétés.
Au lieu de l’atropine, instillons dans l’œil quelques gouttes du poison antagoniste, et c’est alors le sympathique en tant que nerf inhibiteur du sphincter irien et aussi du muscle ciliaire qui sera paralysé. L’ésérine ou la nicotine aurait un effet du même genre encore bien plus marqué. Je n’ai pas à revenir sur ce que j’ai dit plus haut de la double action parallèle, mais fort inégale, d’un même poison sur les deux nerfs antagonistes. Ces vérifications ont toutes été faites avec soin, toutes elles nous amènent à cette conclusion qui, elle aussi, a bien son importance : Ces diverses substances, qui sont des poisons curarisants d’un ordre spécial, se rapprochent du curare encore en ceci qu’ils n’agissent qu’autant qu’ils atteignent les nerfs à leur périphérie et non à leurs origines dans les centres encéphalo-médullaires.
V.
Mais ici une question se pose à nous : Que faut-il entendre par ces mots : « les extrémités terminales » des nerfs moteurs ? Il faut distinguer s’il s’agit d’un nerf ordinaire, la réponse est facile. Un tel nerf, je veux dire la fibre nerveuse, qui en est l’élément constituant, est pour nous un organe simple, manifestant les mêmes propriétés, aussi bien qu’il garde la même structure depuis ses origines dans la moelle jusqu’à sa terminaison dans le muscle : c’est à cet élément à fonction d’ordre volontaire que le curare s’adresse de préférence. Rien de plus facile que de dire où est exactement son extrémité. Mais s’il s’agit, au contraire, des nerfs qui affectent les poisons qui nous occupent, c’est tout autre chose : le sympathique n’est pas un nerf simple ; le pneumogastrique non plus, ni l’oculo-moteur, au moins en tant qu’ils commandent l’un à la pupille, l’autre au cœur ; ce sont des nerfs moteurs ganglionnaires, et il me paraît du plus haut intérêt de voir que l’analyse toxicologique des propriétés de ces nerfs les classe précisément comme nous les classons d’autre part d’après leurs caractères tirés tant de l’anatomie que de la physiologie. C’est une preuve, sans doute, qu’ils constituent les membres épars d’un même système, le système nerveux de la vie involontaire par opposition aux nerfs moteurs non ganglionnaires qui transmettent aux muscles du squelette les ordres de la volonté.
Par ce fait que sur son trajet depuis la moelle jusqu’aux organes la fibre sympathique se met en relation avec une ou plusieurs cellules ganglionnaires, elle n’a plus seulement une extrémité terminale, elle en a nécessairement plusieurs ; elle en a autant qu’il y a de segments inter-ganglionnaires entre la moelle d’une part et le muscle ou la glande d’autre part. Laquelle de ces extrémités agit le poison, je veux dire à quel niveau se fait sa pénétration ? Il est actuellement bien difficile de répondre à une pareille question. Tout ce que je puis affirmer, c’est que ces substances toxiques n’affectent point l’extrémité du nerf qui tient à la moelle, autrement dit ses origines ; et dans la plupart des cas leur point de pénétration est voisin de la périphérie, témoin les expériences sur la pupille et sur les glandes sudoripares.
Cette question du rôle des cellules ganglionnaires et des l’apport qui existent entre ces éléments et les fibres du nerf qui traversent les ganglions est d’une grande importance et d’un grand intérêt pour la compréhension générale du système nerveux, et son étude a été abordée déjà à divers points de vue. (Dastre et Morat, Archives de physiologie ; Langley et Dikinson, Journal of Physiology ; Langendorff, Centralblatt für Physiologie.)
Que l’on comprenne cependant bien ma pensée : je ne prétends nullement dire que les deux poisons dessus désignés portent leur action sur les cellules nerveuses du grand sympathique, sur ses ganglions terminaux ou autres ; par raison d’analogie, je ne le pense même pas. Pour ce qui est du curare, par exemple, il est prouvé jusqu’à l’évidence que ce poison ne s’adresse pas aux cellules d’où émanent les nerfs moteurs, mais tout au contraire aux fibres nerveuses elles-mêmes, à l’exclusion des cellules, ou peut-être même, comme le pensait Vulpian, à quelque organe spécial intermédiaire au muscle et au nerf (plaque motrice) ; il est difficile de décider sur ce dernier point, mais, en tout cas, ce qui est affecté par le toxique, ce qui est paralysé en un mot, c’est le nerf et non le centre. On admettra qu’il y a des raisons de supposer que pour l’atropine et la pilocarpine il en est de même ; leur action, sans doute, porte sur la fibre intra ou inter-ganglionnaire et non sur le ganglion.
VI.
Le grand sympathique, soit seul, soit concurremment avec le pneumogastrique, innerve encore bien d’autres organes que ceux ci-dessus désignés (l’iris, le cœur et les glandes). Ces deux nerfs gouvernent, entre autres, les mouvements de l’estomac, et là, comme pour le cœur, ils représentent deux puissances antagonistes, mais inversées dans leur fonction, en ce sens que c’est le vague qui est excitateur et le sympathique qui est inhibiteur de ces mouvements (von Brani-Hongkest, Morat). On sait depuis longtemps aussi que le sympathique exerce une action d’arrêt sur l’intestin (Pflüger). Il y avait donc lieu de poursuivre sur ce terrain l’analyse commencée plus haut. J’ai montré depuis longtemps, dans mes cours, combien une telle recherche est rendue précise et commode par l’application de la méthode graphique. On introduit dans l’estomac par la bouche et l’œsophage une ampoule élastique, que l’on distend ensuite modérément en y introduisant de l’air ou de l’eau. Cette ampoule, maintenue appliquée contre la paroi gastrique par une contre-pression extérieure, est mise en communication avec un tambour de Marey, dont le style enregistre les contractions de cette paroi sur un cylindre tournant.
L’injection d’un à deux centigrammes de pilocarpine dans le sang de l’animal se traduit par une exagération considérable des mouvements de l’estomac ainsi que de son tonus musculaire. La méthode employée est peu propre à mettre en évidence le péristaltisme de ces contractions, mais elle montre bien leur caractère rythmique, trait commun à un grand nombre des mouvements de la vie involontaire. Pendant que ces contractions se succèdent ainsi avec une grande activité (ils peuvent durer pendant des heures), vient-on à injecter dans la veine moins d’un milligramme d’atropine, on les voit tout d’un coup cesser et l’estomac rester comme paralysé. Si on découvre alors le vague au cou pour l’exciter, on voit qu’il a perdu son excitabilité motrice à l’égard de l’estomac, ses éléments gastro- moteurs sont paralysés, comme le sont du même coup ses éléments cardio-inhibiteurs. Si, avant l’injection d’atropine et pendant que l’animal était encore sous l’influence de la pilocarpine, on excite le tronc du splanchnique (nerf inhibiteur de l’estomac comme de l’intestin), on voit que l’excitabilité de ce tronc nerveux a également diminué ou disparu suivant la dose employée. Là encore les deux substances agissent comme des curares s’adressant d’une façon élective l’un à l’un et l’autre à l’autre des deux systèmes nerveux opposés qui règlent les contractions gastriques. Et en disant que c’est une action élective, je fais la même réserve que plus haut ; cette action, en apparence spécifique, est simplement prédominante tantôt sur l’un tantôt sur l’autre des deux systèmes.
Le même antagonisme peut s’observer pour les mouvements de l’intestin et s’interpréter de même sur ce dernier viscère. J.A. Prévost l’avait observé déjà entre l’atropine et la muscarine, un poison dont l’action est très semblable à celle de la pilocarpine. J’ai de même appliqué la méthode graphique à l’étude des contractions de la vessie urinaire, et soumis dans ces conditions l’animal à la double intoxication successive par les deux poisons : la pilocarpine provoque des contractions très énergiques des muscles vésicaux, et si l’on empêche ces contractions d’avoir leur effet physiologique habituel, l’expulsion de l’urine par le col vésical, on les voit se produire avec un caractère un peu imprévu. Elles sont rythmiques et, à la durée près, elles ressemblent assez à celles de l’estomac ou de l’intestin. J’ai encore examiné l’influence des deux mêmes agents sur les uretères dont la vessie n’est, en somme, qu’une portion dilatée et transformée en réservoir. Ces canaux, dont la fonction est d’assurer l’écoulement continu de l’urine du rein à la vessie, aident en réalité à cette progression par des mouvements propres qui ne sont pas sans analogie avec ceux de l’œsophage et de l’intestin. Lorsqu’à travers une plaie lombaire ou abdominale on les a mis à découvert, on les voit présenter, à des intervalles réguliers, des mouvements vermiculaires, péristaltiques, dirigés du rein à la vessie, chacune de ces ondes de contraction poussant devant elle une onde liquide dans la même direction. Ces mouvements sont exagérés et accélérés par la pilocarpine ; ils sont diminués et retardés (je ne suis pas arrivé à les supprimer) par l’atropine. L’utérus fait encore de même lorsque sa fonction physiologique l’a mis en état de se contracter, c’est dire vers la fin de la gestation ; la pilocarpine provoque et accélère les crampes expulsives dont il est alors le siège, et l’atropine les retarde ou les fait cesser [3].
En somme, comme on voit, le double effet de ces substances tend à se généraliser de plus en plus à toute une classe d’organes entre lesquels l’analyse physiologique décèle une parenté évidente, surtout en ce qui concerne leurs relations avec le système nerveux.
VII.
Mais le grand sympathique étend encore son influence à la fois excitatrice et inhibitrice sur d’autres organes encore plus intimement liés, si possible, que les précédents, à la vie involontaire ou de nutrition, les vaisseaux. Quelle est donc l’action de la pilocarpine et de l’atropine sur le système vaso-moteur ?
Cette action est indéniable, évidente. Elle a, du reste, été déjà signalée maintes fois, tant par les cliniciens que par les physiologistes. Elle a été étudiée à différents points de vue et à l’aide de méthodes variées (Mosso). Mais je m’empresse d’ajouter que cette action n’est pas aussi univoque, aussi catégorique que celle observée sur les autres nerfs moteurs et inhibiteurs du grand sympathique. Tandis que pour ceux-ci la paralysie survient pour des doses parfois infinitésimales, il faut pour les vaso-moteurs employer des doses bien plus fortes, si on veut observer une modification appréciable de l’excitabilité de ces nerfs. Enfin l’action antagoniste ne s’y montre pas non plus aussi évidente que dans les exemples précédents. Il ne faut pas oublier, du reste, que l’ensemble du système vasculaire se décompose en départements ou sous-systèmes qui, sous des influences en apparence identiques, réagissent souvent à l’inverse les uns des autres. La seule étude des poisons vaso-moteurs nécessiterait une analyse très longue et détaillée dans laquelle chacun des organes principaux devrait être interrogé d’une façon spéciale. Je ne puis me dissimuler qu’une telle étude présentera encore des lacunes et nécessitera souvent des retouches. Toutefois, les faits observés jusqu’ici me conduisent une fois de plus à la même conclusion : 1° les deux poisons examinés agissent en paralysant plus ou moins les nerfs vaso-moteurs, et cette inégalité dans la paralysie de deux ordres de nerfs, l’un dilatateur, l’autre constricteur, détermine le sens de la réaction apparente du système vasculaire ; 2° les modifications d’excitabilité de nerfs vaso-moteurs, bien que moindres (aux mêmes doses) que celles des autres nerfs centrifuges involontaires, sont pourtant encore bien plus appréciables que celles des nerfs moteurs volontaires, en entendant par cette expression la portion du système moteur qui est en dehors de la cavité encéphalorachidienne, c’est-à-dire en faisant abstraction de toute la portion intra-osseuse du système nerveux qu’on appelle communément les centres. La comparaison entre les deux systèmes moteurs, volontaire et involontaire, ne pouvait bien se faire, au point de vue expérimental, que dans ce qu’on appelle communément le système nerveux périphérique.
Notre affirmation du début se trouve donc ainsi légitimée : les substances sont avant tout des poisons du système sympathique. Mais qu’est-ce que le système sympathique ?
VIII.
L’idée qu’un ensemble de nerfs distincts des autres nerfs est attribué au gouvernement des fonctions de nutrition est ancienne en médecine, pour ne pas dire en physiologie. Cette idée y a été, sinon introduite, du moins développée et défendue par Bichat, qui en fait la base de sa conception du système nerveux. Elle a été évidemment suggérée par l’anatomie. La pensée ne viendrait en effet pas à un anatomiste de décrire séparément les différentes branches du sympathique en les intercalant parmi les nerfs qui font suite aux paires nerveuses sortant de la colonne par les trous de conjugaison. Tous les anatomistes ont toujours décrit le sympathique comme un système à part, soit qu’ils lui donnent ou qu’ils lui refusent cette désignation. Les rapports du sympathique avec la nutrition ont donc été pressentis avant que I’on connût, soit les fonctions de ce nerf, soit le peu que l’on sait encore des actes de la nutrition. En un mot, c’est une vue synthétique, une induction qui s’est trouvée juste.
La. physiologie a dû procéder par une voie inverse, c’est-à-dire par l’analyse expérimentale, et quand elle a eu recueilli de-ci de-là quelques exemples épars de ces nerfs qui président à des actes involontaires, les rapports de la nutrition avec le système nerveux lui sont d’abord apparus tout autrement. Ces données disparates et incomplètes ne se prêtaient pas à une classification. La conception de Bichat fut abandonnée par les physiologistes, et le sympathique ne fut d’abord pour eux qu’un nerf quelconque semblable à tous les autres.
Les choses en étaient là quand, Dastre et moi, nous avons fait connaître un fait qui était de nature à changer de nouveau les idées des physiologistes sur la constitution du grand sympathique comme sur ses rapports avec la circulation en partant avec la nutrition. Ce fait est l’existence, dans le sympathique, d’éléments inhibiteurs des muscles vasculaires, autrement dit de nerfs vaso-dilatateurs juxtaposés pour ainsi dire aux éléments moteurs de ces mêmes vaisseaux. La coexistence dans le même tronc de deux ordres de nerfs, les uns excitateurs, les autres inhibiteurs, me parait tout à fait caractéristique ; c’est le type, en effet, auquel on tend de plus en plus à ramener tout système moteur. Ainsi le système vaso-moteur, disloqué auparavant entre tous les rameaux nerveux, se trouve ramené à l’unité (en tant que système) ; les cieux pièces essentielles qui le composent entrent, en effet, en connexion étroite au niveau des ganglions, soit de la chaine, soit de la périphérie. Nous avons signalé, Dastre et moi, plusieurs exemples bien remarquables de cette fonction à la fois tonique et inhibitoire des ganglions du grand sympathique. Les travaux poursuivis depuis, soit par nous-même, soit par d’autres, sont tout à fait confirmatifs d’une telle vue : ils établissent de plus en plus l’existence pour chaque ordre de mouvements ou de fonction gouvernés par le sympathique de deux ordres de fibres antagonistes et aussi le rôle des cellules ganglionnaires comme point de raccordement de ces deux ordres d’éléments, pour leur permettre d’agir l’un sur l’autre et équilibrer de la façon la plus parfaite les mouvements qu’ils doivent régir.
Mais si la conception de Bichat se trouve vérifiée par ces travaux, il est presque superflu de dire qu’il a fallu la modifier et l’étendre pour l’adapter aux faits nouveaux. Lorsque la même idée fait en science plusieurs apparitions, c’est rarement sous la même forme qu’elle se présente, d’où la difficulté qu’elle éprouve à se faire bien comprendre. Si l’on se place au point de vue très simple de Bichat, les objections tirées de la physiologie ne manquent pas : c’est d’abord celle-ci. Le grand sympathique, c’est vrai, ne contient que des nerfs involontaires ; mais l’inverse n’est pas exact. Les nerfs crâniens contiennent presque tous des éléments à fonction involontaire, témoins le trijumeau et l’oculomoteur : il est facile de répondre à cet argument.
J’ai montré comment la chaine même du sympathique se continue dans le crâne ; elle est représentée par l’anastomose (assez volumineuse chez les animaux) étendue entre le ganglion cervical supérieur et le ganglion de Gasser, laquelle contient des nerfs vasomoteurs, sécréteurs, accommodateurs, irido-dilatateurs, etc. Après avoir traversé le ganglion de Gasser, elle se répartit entre les ganglions ophtalmique, sphéno-palatin et otique, qui sont évidemment des ganglions du sympathique. Au niveau de chacun d’eux, cette portion crânienne de la chaîne se comporte comme au niveau de ses autres ganglions, c’est-à-dire elle entre en rapport avec les paires nerveuses voisines pour leur céder des nerfs (rameaux de distribution) et pour en recevoir (rameaux d’origine). Le trijumeau (en lui adjoignant les nerfs moteurs de l’œil) est justement celui qui rappelle le mieux la disposition typique des paires nerveuses rachidiennes. Seulement dans le crime la simplicité schématique qui se trouve au rachis a disparu, et il faut faire quelque effort d’analyse pour retrouver les portions équivalentes. Au lieu de simples numéros d’ordre, premier, deuxième, etc., les nerfs ont des noms propres (moteur commun, trijumeau), et c’est de cette circonstance grossière que sont nées les objections.
Mais pour les nerfs crâniens, la région la plus bouleversée, c’est celle qui répond à la partie inférieure du bulbe d’où naissent le glosso-pharyngien, le vague, le facial. Là tout est ou paraît confondu : nerfs moteurs, sensitifs, sympathiques. Les caractères purement descriptifs ont disparu, mais la physiologie sait distinguer ces nerfs de fonctions différentes, et dans le vague, le facial, le glosso-pharyngien, elle retrouve des éléments équivalents de ceux du grand sympathique.
Les caractères anatomiques autres que ceux purement descriptifs se retrouvent. Ces nerfs contiennent de fines fibres myéliniques accompagnées de fibres de Remak, et ces fibres sont en rapport avec des ganglions. C’est ce que nous voyons dans le vague, la corde du tympan, les nerfs pétreux, y compris ceux qui viennent du rameau de Jacobson et qui ont tous les caractères structuraux des nerfs sympathiques. Ils naissent dans le bulbe d’une région de substance grise qui, aux yeux des anatomistes, représente évidemment la continuation des origines médullaires du sympathique. Ils aboutissent à des masses ganglionnaires de nature aussi évidemment sympathique : il leur manque ce seul caractère de ne pas faire partie de la chaîne qu’ils croisent sans y entrer, et encore voyons-nous chez beaucoup d’animaux, en particulier chez le chien, le vague s’unir étroitement au sympathique dans la région cervicale, au point de ne pouvoir pas en être dissocié. La première et la plus ancienne classification des nerfs, au temps de Magendie et de Ch. Bell, a du reste rencontré dans cette même région les mêmes difficultés : le nerf bulbaire, qu’on désigne volontiers sous le nom de racine antérieure ou postérieure de tel autre, ne doit point cette désignation à sa position, mais à l’analogie de ses fonctions avec celles des racines médullaires, et ce langage ne cesse pas d’être clair pour être au fond inexact.
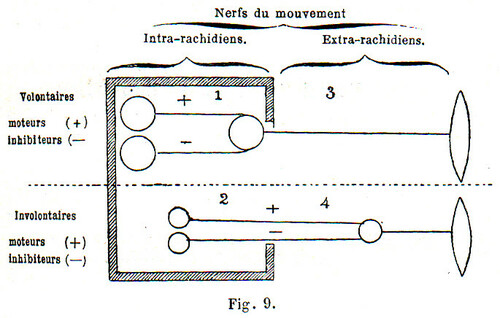
Il n’est pas de systématisation plus simple que la nôtre et, croyons-nous, plus justifiable ; il n’en est pas qui ait été plus mal comprise : il faut dire aussi que les anciennes désignations en usage sont de nature à jeter sur cette classification la plus grande confusion. Le terme de « système cérébro-spinal » opposé à celui de « grand sympathique » n’a pas de sens, outre qu’il constitue une erreur, car il laisse supposer que le partage des deux systèmes, l’un volontaire, l’autre involontaire, se fait entre la chaîne et la moelle, ce qui est inexact : il y a une partie de la moelle et des centres qui appartient au sympathique. C’est donc encore là une donnée de l’anatomie descriptive mal interprétée, qui a conduit à la confusion. Le schéma suivant me paraît représenter clairement la conception qu’on doit se faire du partage des nerfs centrifuges suivant la nature de leurs fonctions (volontaire ou involontaire), le sens de leur action (positive (+) ou négative (-) et en tenant compte de leur caractéristique anatomique principale d’avoir leurs centres immédiats ou distaux, en dehors ou en dedans de la cavité encéphalo-rachidienne. Les divisions tracées dans le sens vertical sont d’ordre anatomique ; celles tracées horizontalement sont, de plus, d’ordre physiologique. Les numéros 1, 2, 3 correspondent à ce qu’on appelait autrefois le système cérébro-spinal. Pour que cette expression ait un sens, il faudrait le réduire aux numéros 1 et 2, et il équivaudrait, par opposition aux nerfs périphériques, au mot de centres usité en anatomie, mais qui n’est qu’une métaphore, attendu que d’autres centres existent en dehors du rachis et du crâne, et que ces centres eux-mêmes sont accompagnés d’une masse énorme de substance blanche représentant des systèmes conc1ucteurs. Le numéro 4 correspond à l’ancien sympathique isolé de ses centres profonds ou intra-rachidiens ; il faut lui adjoindre le numéro 2, et il représente alors le système moteur involontaire par opposition avec les numéros 1 et 3 auxquels certains donnent encore le nom de système cérébro-spinal, en prenant ce mot dans son acception physiologique comme synonyme de système nerveux volontaire [4].
IX.
Il est certainement d’un haut intérêt de voir des substances comme celles que nous avons nommées plus haut agit d’une façon élective sur tel ou tel de ces groupements nerveux et à l’inverse de ce qu’on suppose souvent devoir être et de ce qui se remarque avec d’autres poisons ; de voir ces poisons atteindre le système nerveux par ses parties les moins élevées et respecter d’abord les autres ; de les voir distinguer, même en lui, non seulement les systèmes, mais les systèmes dont il est formé, et déséquilibrer les fonctions organiques à la volonté de l’expérimentateur. Mais, sur ce point justement, je voudrais qu’on ne se méprenne pas sur ma pensée. En les appelant les curares du grand sympathique, je n’entends nullement dire que leur action y reste confinée. Cette action est bien plus générale et plus profonde : on la voit s’étendre peu à peu aux nerfs du mouvement volontaire ; mais, chose bien digne de remarque, elle reste localisée à certaines parties de ce système situées en amont des centres d’origine des nerfs moteurs, de telle façon que ceux-ci, interrogés directement, paraissent indemnes. C’est en cela surtout que réside la différence des effets plus apparente que réelle et rendue si frappante seulement par la juxtaposition dans l’ensemble des nerfs périphériques de parties non équivalentes réagissant chacune à sa façon, suivant le type structural ou fonctionnel auquel elle appartient.
En d’autres mots : dans le domaine de la vie volontaire, ces poisons s’adressent aux éléments intra-rachidiens ; dans le domaine de la vie involontaire, ils s’adressent de plus à des éléments extra-rachidiens. Pourquoi ? Parce que dans le premier les puissances antagonistes sont confinées en dedans du crâne et du rachis, tandis que dans le second elles se prolongent en dehors, comme le schéma ci-dessus le fait comprendre.
Ce qui est curieux dans l’analyse de ces effets de l’atropine et de la pilocarpine sur toutes les grandes fonctions, c’est de voir leur antagonisme se poursuivre, se confirmer à mesure qu’on les étudie davantage. J’ai montré depuis longtemps que l’atropine abaisse la proportion de sucre contenu dans le sang, tandis que la pilocarpine l’élève faiblement, il est vrai, mais d’une façon constante. Ces variations se lient certainement à une consommation différente du sucre du sang dans les deux cas plus encore qu’à des variations dans sa production. Chez l’animal intoxiqué par l’atropine, on voit la température s’élever et rester plusieurs jours au-dessus de la normale. De plus sa respiration s’accélère : l’animal est éveillé, très excitable, et montre une grande tendance au mouvement. C’est l’inverse avec la pilocarpine : la respiration est lente, l’animal est déprimé et sa température s’abaisse. Bien certainement les deux substances atteignent là encore, dans les centres mêmes de la vie de relation ; deux ordres de nerfs antagonistes, et arrivent de cette façon à déséquilibrer la fonction du mouvement et par contrecoup la respiration, la calorification et la glycémie. Cliniquement, on sait que l’intoxication par la belladone élève la température et, outre le délire, produit parfois des convulsions.
X.
Dans la question encore controversée de savoir si l’antagonisme entre l’atropine et la pilocarpine est réel ou, comme on dit, réversible et bilatéral, on peut voir par ce qui précède quelle position je prends parmi ceux qui l’ont discutée. Mon opinion se rapproche et s’éloigne à la fois de chacune de celles défendues jusqu’ici, et je la formule de la façon suivante : « Oui, il y a dans de certaines limites antagonisme réel, réversible, bilatéral, c’est démontré ; seulement cet antagonisme n’est pas entre les substances elles-mêmes, mais entre les éléments constituants du système de nerfs qui régit la fonction influencée. Les deux substances et leurs succédanées ont une action paralysante analogue à celle du curare ; cette action porte inégalement sur les deux ordres d’éléments, soit moteurs, soit inhibiteurs, qu’on démontre exister dans les systèmes particuliers qui régissent les mouvements involontaires ou de la vie végétative. L’étroitesse des limites entre lesquelles s’observe la réversibilité s’explique justement par l’action inégalement paralysante, inversement paralysante, mais dans les deux cas paralysante des deux agents. Si l’antagonisme était entre les substances elles-mêmes, il serait indéfini et non limité. Le fait que la même substance déséquilibre les différentes fonctions au profit, tantôt de l’élément moteur, tantôt de l’élément inhibiteur, s’explique également mieux par une action paralysante d’ordre un peu général portant sur les deux à la fois, mais inégalement. De même que le curare, ces poisons atteignent les nerfs pal’ leur périphérie et non par leurs origines. Mais à l’inverse du curare, ils s’adressent aux segments inter-ganglionnaires (fibres inter-centrales) et non aux segments terminaux : du système ; à l’inverse également du curare, ils affectent primitivement les nerfs de la vie involontaire, puis à dose croissante les nerfs de la vie de relation. C’est en tenant formellement compte de cette restriction qu’on peut les appeler, comme je l’ai fait, les curares de la vie végétative, comme le curare lui-même est le poison paralysant par excellence des nerfs volontaires. L’effet antagoniste de ces substances, constaté également dans le champ d’action des nerfs volontaires, s’ajoute aux présomptions, on peut même dire, dès aujourd’hui, aux preuves de l’existence dans le système volontaire de deux’ ordres d’éléments, les uns moteurs, les autres inhibiteurs. Chacune de ces substances a des succédanées qui reproduisent ses propres effets dans leurs grands traits, mais qui se distinguent d’elle par quelque nuance, ne fût-ce que quantitativement. L’étude de toutes ces différences est du plus grand intérêt pour le médecin qui en peut tirer des effets thérapeutiques divers, pour le physiologiste qui trouve dans ces substances des agents extrêmement précieux et, j’ose dire, pour le psychologue qui peut, lui aussi, à l’occasion, y trouver des procédés d’analyse.
J.-P. MORAT.
