Dans un précédent article [1], j’ai montré qu’on pouvait saisir par la photographie les attitudes successives des ailes d’un oiseau à différents instants de son vol ; qu’on obtenait en même temps les positions de l’oiseau dan l’espace à des intervalles de temps égaux entre eux et parfaitement connus, et j’émettais alors l’espérance de résoudre par cette méthode le problème si obscur du mécanisme du vol.
Depuis lors, la méthode s’est perfectionnée et le nombre des espèces d’oiseaux sur lesquelles ont porté mes études s’est multiplié.
De la comparaison des différentes espèces que j’ai eues à ma disposition, il résulte que, sauf certaines différences de détails, tous les oiseaux exécutent des mouvements de même nature : chez tous, en haut de sa course, elle se déploie brusquement l’aile se replie au moment de sa remontée ; arrivée puis s’abaisse en se portant en avant et en se rapprochant du corps ; à la fin de cet abaissement, les articulations des ailes se replient de nouveau et la montée recommence.



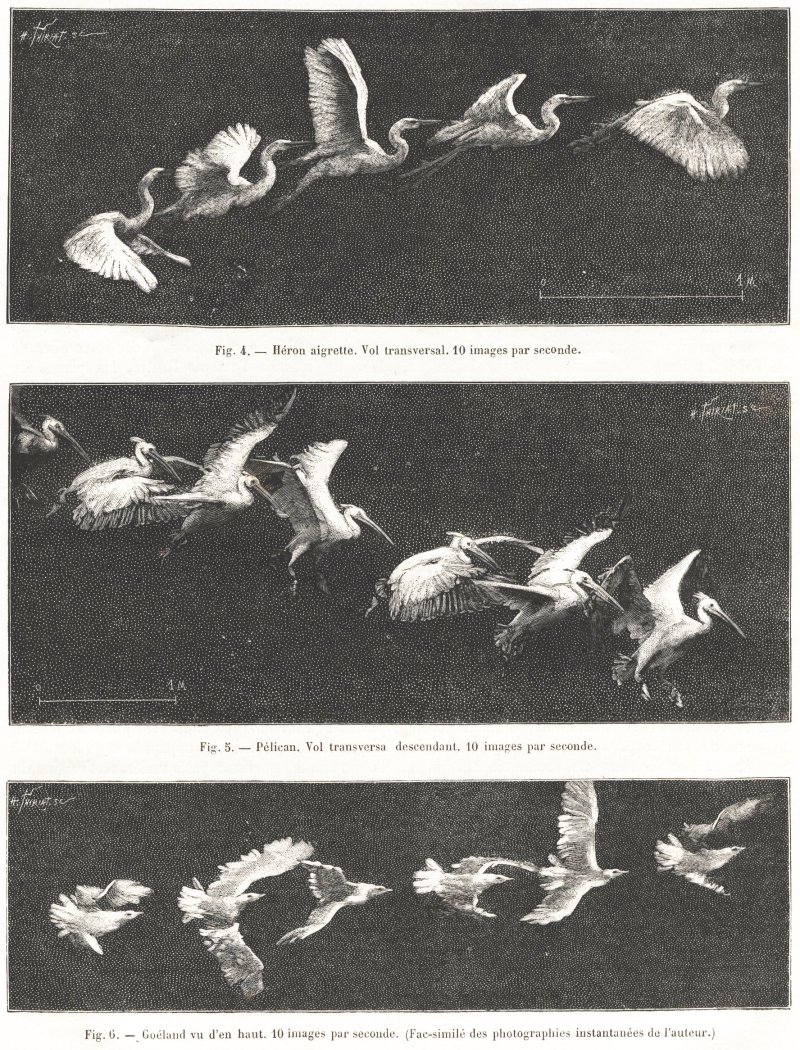
Les figures 1, 2, 3, 4 et 5 représentent le vol du goéland, du héron, du pigeon et du pélican.
Ces figures révèlent de curieuses attitudes que l’œil n’a pas le temps de saisir et avec lesquelles nous ne sommes point familiarisées par les représentations artistiques des oiseaux. Suivant une juste remarque de M. Muybridge, les peintres européens représentent, à peu près toujours, les oiseaux au vol avec les ailes levées ; les Chinois et les Japonais, au contraire, les représentent indifféremment dans l’élévation ou dans l’abaissement des ailes. Ce n’est pas à dire pour cela que les artistes de l’extrême Orient aient fidèlement reproduit les différentes attitudes des oiseaux : la comparaison de leurs images avec celles que donne la photographie instantanée montre bien que, pas plus en Chine que chez nous, l’œil humain ne peut surprendre des actes qui ne durent qu’un très court instant.
Vues sous un seul aspect, les images d’un oiseau qui vole ne renseignent pas encore suffisamment sur la nature des mouvements des ailes ; il faut photographier l’oiseau sous plusieurs aspects différents pour bien comprendre ce mécanisme. Nous avons disposé plusieurs appareils à cet effet. L’un, placé à 12 mètres de hauteur, donnait des images de l’oiseau vu d’en haut (fig. 6) d’autres orientés différemment le montraient de côté, ou bien volant dans la direction de l’objectif photographique (fig. 7). Ces images prises sous différentes incidences se complètent l’une par l’autre. Ainsi, les oiseaux vus d’en haut montrent une singulière torsion du plan de l’aile dont on ne soupçonnerait pas l’existence d’après les images prises latéralement. Cette torsion se produit à la fin de l’abaissement de l’aile, au moment où les articulations commencent à se plier pour préparer la remontée. Il en résulte un aspect hélicoïdal de l’aile rappelant la forme que M. Pettigrew considère comme la cause essentielle de la pro pulsion de l’oiseau. Mais il faut observer que cette forme ne se produit qu’à la lin de l’acte d’abaissement, au point mort de Faction de l’aile, comme on dit en mécanique, et dans un moment où celle-ci, devenue passive, va être remontée par la résistance de l’air. Ces images montrent encore un fait assez imprévu, c’est que les actes du vol ne sont point symétriques. On avait déjà supposé avec raison que l’oiseau qui veut infléchir latéralement la direction de son vol exécute des mouvements plus étendus du côté qui doit progresser le plus vite ; c’est-à-dire qu’il donne plus d’amplitude aux mouvements de l’aile droite s’il veut tourner à gauche et réciproque ment. Il est à peine nécessaire de dire que la photo chronographie condamne entièrement l’hypothèse dans laquelle on supposait qu’une des ailes de l’oiseau pouvait battre plus fréquemment que l’autre ; les mouvements des deux ailes sont parfaitement synchrones, sinon égaux en étendue. On voit enfin sur ces images, que le corps de l’oiseau s’incline et se tord de diverses manières, de façon à porter son centre de gravité d’un côté ou de l’autre, suivant les besoins de l’équilibre. L’oiseau dont les attitudes sont représentées (fig. 6) semblait sollicité à porter le poids de son corps à gauche, par suite de la moindre surface de son aile droite oit manquent quelques pennes.

Quant aux images prises de face et un peu oblique ment comme dans la figure 7, elles renferment aussi d’utiles renseignements. Elles font voir que l’extrémité de l’aile, partie réellement active de l’organe, puisqu’elle frappe l’air avec plus de vitesse, présente, au moment de l’abaissement, des changements de plan que n’offrent pas les rémiges secondaires qui s’étendent du carpe à l’épaule. Il se fait, entre les pennes des différents ordres, une sorte de séparation qui montre que l’articulation du carpe est le siège d’un léger mouvement de torsion favorable à l’inclinaison du plan des rémiges carpiennes. Sur ces images on voit encore très bien la courbure et la convergence des ailes à la lin de leur abaissement, l’encoche que présente à ce moment le bord antérieur de l’aile, par l’effet d’un commencement de flexion du coude. On conçoit que pour suivre dans tous leurs détails les changements d’attitudes des ailes, il ait fallu multiplier beaucoup les expériences, de manière à obtenir, pendant un seul coup d’aile, dix ou douze attitudes successives de l’oiseau vu sous chacun de ses différents aspects.
Ces images une fois obtenues, j’étais en possession de tous les éléments nécessaires pour comprendre complètement les mouvements des ailes suivant les trois dimensions de l’espace. Mais, pour les représenter, des ligures en relief devenaient nécessaires ; les circonstances me servirent à souhait.
A Naples, où je me trouvais alors, l’industrie de la fonte du bronze à cire perdue s’est conservée depuis la plus haute antiquité. Je modelai en cire une série de figures représentant les attitudes successives dans une même révolution de l’aile, dix images pour le goéland, onze pour le pigeon ; ces maquettes livrées à un habile fondeur furent reproduites en bronze avec une fidélité parfaite.
La figure 8 représente, disposées en série et suivant leur ordre de succession, à des intervalles de 1/88 de seconde l’une de l’autre, les phases du coup d’aile d’un pigeon.

Pour les besoins de la photographie on a peint en blanc ces figures de bronze, afin d’y rendre plus sensibles les effets d’ombre et de lumière. Grâce à la multiplicité des attitudes représentées dans cette série, on y suit facilement toutes les phases du mouvement des ailes ; on voit comment elles se replient, s’élèvent, se déploient et s’abaissent.
Pour mieux faire comprendre comment s’enchaînent entre eux les mouvements de l’aile de l’oiseau dont la photochronographie a ’donné l’analyse, j’ai recouru à l’emploi du zootrope qui les recompose et rend à la vue l’impression que donne un oiseau qui vole.
Le zootrope représenté ci-après (fig. 9) offre cette particularité, qu’il est formé par des figures en relief. C’est un grand avantage au point de vue de l’impression qu’il donne ; en effet, ces figurines d’oiseau, disposées en cercle dans l’appareil, se présentent à l’observateur sous des aspects variés. A l’origine du mouvement, les oiseaux sont vus de dos ; puis, dans leur trajet circulaire, ils présentent leur flanc, passent en plein travers, et reviennent enfin sur l’observateur. En outre, les mouvements de l’aile qui, dans la nature, sont extrêmement rapides et par conséquent insaisissables, sont ici beaucoup ralentis, de sorte qu’on en peut suivre aisément les phases et se rendre compte en un instant de ce que l’observation la plus attentive du vol des oiseaux ne permettrait pas de saisir. La figure 9 montre la disposition du zootrope ; elle ne peut malheureuse ment pas donner une idée de l’effet produit par l’ap pareil en mouvement.

Mais, dira-t-on, cette méthode cinématique traduit les mouvements de l’oiseau sans indiquer la force qui les produit. Il serait pourtant bien utile de connaître cette force, mieux encore de mesurer le travail mécanique dépensé par l’oiseau pour se soutenir et se transporter dans les airs.
Voyons ce que nos images photographiques nous révèlent à cet égard.
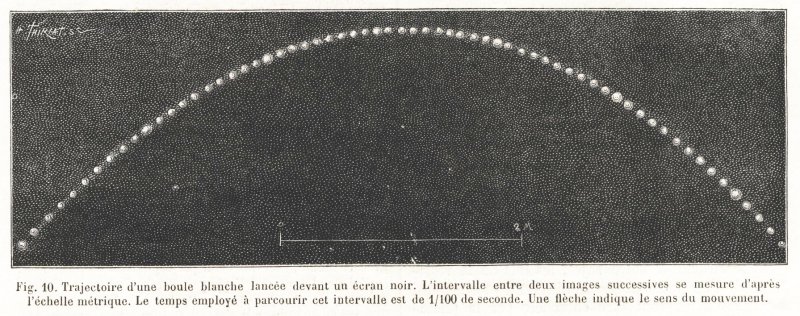
Quand on connaît la masse d’un corps et la vitesse dont il est animé, on connaît, par cela même, la force qui a mis ce corps en mouvement et le travail dé pensé par cette force. Soit un projectile d’un poids connu, nous le lançons devant l’appareil photochronographique et nous prenons une série d’images de ce projectile à des intervalles de 1/100 de seconde. La figure 10 montre la trajectoire suivie, et l’écartement qui sépare les images les unes des autres fait connaître l’espace parcouru par le projectile dans chacun des centièmes de seconde qu’a duré son mouvement. De dix en dix, une image plus brillante a été produite par une fenêtre du disque plus large que les autres ; ces repères sont utiles pour faci liter la numération des images. Une échelle métrique fixe, photographiée en même temps que l’objet en mouve ment, sert à mesurer les espaces parcourus à chaque instant. Voilà donc un problème de balistique dont la solution peut être facile-nient obtenue par les méthodes usuelles de calcul.
Les images successives de l’oiseau qui vole se prêtent à la même analyse dynamique. La balance, en nous indiquant le poids de l’oiseau, nous en fera connaître la masse. Et pour que la photochronographie nous donne avec perfection la trajectoire de cette niasse, il suffira de multiplier beaucoup les images recueillies ; on en prendra cent par seconde au besoin. Mais alors ces images se confondront partiellement entre elles, car l’oiseau, en un centième de seconde, ne parcourt pas un espace égal à la longueur de son corps ; il en résultera que la seconde image se superposera en partie à la première, la troisième à la seconde, et ainsi de suite. A peine, dans cette confusion, pourra-t-on distinguer le moment où l’aile s’abaisse de celui où elle s’élève. Qu’importe : nous fixerons sur la tête de l’oiseau un petit point métallique très brillant et l’image de ce point, nettement visible dans la série des figures, révélera la trajectoire de l’oiseau avec sa vitesse et avec les accélérations et les ralentissements que produisent les mouvements des ailes. On peut alors aborder le problème dynamique du vol. On constate d’a bord que l’oiseau n’oscille pas sensiblement dans le sens vertical, d’où l’on doit conclure que la résistance de l’air sous ses ailes est précisément égale à son poids. D’autre part, on observe que la translation de l’animal présente des alternatives d’accélérations et de ralentissements exprimant que la force impulsive ou la résistance de l’air prédomine tour à tour. De la va leur de ces accélérations de signes contraires se déduisent la valeur de la composante horizontale du mouvement de l’oiseau et celle de la résistance de l’air.
Les calculs basés sur ces expériences ont donné les valeurs suivantes pour les forces qui agissent pendant le vol du goéland.
| Composante verticale. | 0kg,623 |
| Composante horizontale. | 0kg,898 |
| Total | 1kg,521 |
Ces forces se développent pendant le coup d’abaissement des ailes quant à la remontée, elle est passive et due à la pression de l’air sous la face inférieure des ailes qui agissent alors pour soutenir l’oiseau à la façon d’un cerf-volant.
Comme la résistance de l’air sous les ailes agit en un point assez éloigné de l’articulation de l’épaule, et comme les muscles pectoraux , abaisseurs des ailes, agissent très près de l’articulation, c’est-à-dire sur un bras de levier plus défavorable, il en résulte que l’effort des muscles est bien plus grand que la résistance de l’air qu’ils surmontent. Pour les pectoraux du goéland l’effort développé serait de 19 kilogrammes.
On s’est demandé souvent si les muscles des oiseaux n’avaient pas une force spécifique plus grande que ceux des autres animaux, c’est-à-dire si deux faisceaux musculaires de même grosseur appartenant, l’un à un oiseau, l’autre à un mammifère, au raient des forces différentes. Sur le goéland qui a servi à mes expériences, une section transversale des muscles pectoraux, pratiquée perpendiculairement à la direction de leurs fibres, avait environ 11 centimètres carrés de surface, soit environ 1 k ,600 par centimètre carré. D’autres oiseaux m’ont donné autrefois des valeurs à peu près semblables pour leur force spécifique ; ainsi la buse développait 1200 grammes par centimètre carré de section musculaire, le pigeon 1400 grammes.
Les adeptes de l’aviation espèrent qu’on réalisera quelque jour une machine capable de transporter l’homme dans les airs, mais beaucoup d’entre eux sont troublés par un doute : ils se demandent si la force de l’oiseau n’excède pas celle des moteurs connus. Les expériences ci-dessus peuvent déjà les rassurer, car si nous comparons la force musculaire des oiseaux à celle de la va peur, nous voyons qu’un muscle serait assimilable à une machine à très basse pression. En effet, la vapeur qui développerait 1k,600 par centimètre carré n’aurait guère plus d’une atmosphère et demie de pression.
Mais la véritable comparaison à établir entre les moteurs animés et les machines, consiste à mesurer le travail que chacun de ces moteurs peut fournir, à poids égal, dans l’unité de temps.
La mesure du travail d’un moteur s’obtient en multipliant l’effort développé, par le chemin que parcourt le point d’application de cet effort. Les photochronographies expriment, à chaque instant, les espaces parcourus par la masse de l’oiseau, et le déplacement du centre de pression de ses ailes, donnant ainsi le facteur chemin dans la mesure du travail. On trouve, d’après cela, que pour les cinq coups d’aile que le goéland donne à chaque seconde, au moment oit il s’envole, le travail effectué serait de 3kg,668. Ce chiffre est très élevé : il correspond à celui que ferait une ma chine en élevant son propre poids à plus de 5 mètres de hauteur par seconde.
Mais ce n’est là qu’un maximum que l’oiseau ne produit qu’au moment de l’essor, lorsqu’il n’a pas encore de vitesse. En effet, à mesure que s’accélère la translation de l’oiseau, l’air présente sous ses ailes un point d’appui plus résistant. J’ai démontré autrefois expérimentalement ce lait déjà annoncé par les frères Planavergne, de Marseille, et dont la théorie est la suivante.
Quand l’oiseau n’a pas encore de translation, l’air qui est frappé par ses ailes présente, au premier instant, une résistance due à son inertie, puis entre en mouvement et fuit au-dessous de l’aile sans lui fournir d’appui. Lorsque l’oiseau est en pleine vitesse, au contraire, son aile s’appuie à chaque instant sur de nouvelles colonnes d’air dont chacune lui offre la résistance initiale due à son inertie. La somme de ces résistances présente à l’aile un point d’appui beaucoup plus ferme. Un pourrait comparer l’oiseau qui s’en vole à un marcheur qui fait de grands efforts pour cheminer sur un sable mouvant et qui, à mesure qu’il avance, rencontre un sol de plus en plus ferme, de sorte qu’il progresse plus vite et avec moins de fatigue. L’accroissement de la résistance de l’air diminue la dépense de travail ; les coups d’aile de l’oiseau de viennent, en effet, moins fréquents et moins étendus. En air calme, un goéland qui a pris sa vitesse dépense à peine le cinquième du travail qu’il devait fournir au début de son vol. L’oiseau qui vole contre le vent se trouve dans des conditions plus favorables encore, puisque des masses d’air qui se renouvellent continuellement apportent sous ses ailes leur résistance d’inertie. C’est donc l’essor qui constitue la phase la plus laborieuse dans le vol. On a remarqué depuis longtemps que les oiseaux emploient toutes sortes d’artifices pour acquérir une vitesse préalable avant de battre des ailes ; les uns courent sur le sol avant de s’élancer dans l’air, ou sautent brusque ment dans la direction qu’ils veulent suivre en volant ; d’autres se laissent tomber d’un lieu élevé les ailes étendues et glissent sur l’air avec une vitesse accélérée avant de battre des ailes ; tous s’orientent le bec au vent au moment de l’essor.

Mes expériences n’ont pu, jusqu’à présent, s’appliquer qu’au vol de départ ; il faut, pour étudier le plein vol, des conditions difficiles à réaliser. Avec une gracieuseté dont je le remercie, M. Eiffel m’a offert, sur la tour gigantesque qu’il érige, un poste d’observation qui ne laissera rien à désirer. De cette énorme hauteur, des oiseaux photographiés pendant un long parcours donneront des images photochronographiques beaucoup plus instructives que celles que j’ai pu obtenir jusqu’ici.
Sans entrer dans les détails arides des expériences et des calculs effectués ( [2]), j’ai voulu montrer que les mouvements de l’oiseau, s’ils échappent à la vue, peuvent être fidèlement traduits par une nouvelle méthode qui s’applique aux problèmes les plus variés de la cinématique et de la mécanique.
La photochronographie, en effet, donne expérimentalement la solution de problèmes souvent très difficiles à résoudre par le calcul. Qu’on imagine un certain nombre de forces agissant en sens divers sur une masse connue ; la façon compliquée dont ces forces se composent entre elles rend parfois de longs cal culs nécessaires pour déterminer les positions que le mobile occupera à des instants successifs ; tandis que si le corps lui-même, soumis à ces différentes forces, peut être placé devant l’appareil photochronographique, la trajectoire qu’il suivra se traduira d’elle-même sur la plaque sensible.
Des physiciens distingués discutaient dernièrement sur la forme que doit présenter l’extrémité libre d’une tige vibrante dans laquelle on produit des ventres et des nœuds ; la plupart d’entre eux supposaient qu’entre le dernier nœud et son extrémité libre la verge présente une forme courbe. L’expérience a. montré qu’il n’en est pas ainsi et que les derniers éléments de la verge vibrante sont parfaitement rectilignes (fig. 11).
Que de problèmes dont la solution a coûté autre fois des efforts de génie, pourraient se résoudre par une expérience fort simple ! Galilée, de nos jours, n’aurait plus besoin de ralentir la chute des corps pour en observer le mouvement. Il laisserait tomber une boule brillante au devant .d’un champ obscur et en recueillerait photographiquement les images successives. Sur la plaque sensible se liraient, le plus simplement du inonde, les lois des espaces, des vitesses et des accélérations qu’il a eu la gloire de découvrir.
Pour revenir à notre sujet, les lois de la résistance de l’air aux mobiles de différentes formes qui s’y meuvent devront être cherchées par la photochronographie. Déjà d’intéressants résultats sont acquis ; nous avons pu déterminer la trajectoire et les vitesses de petits appareils planeurs qui se meuvent librement dans l’air et que l’œil n’a pas le temps de suivre dans leurs mouvements rapides. Ces études reprises et méthodiquement suivies conduiront sûrement à comprendre le mécanisme encore obscur du vol plané.
Étienne.-Jules. MAREY, de l’Institut.
